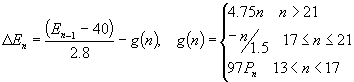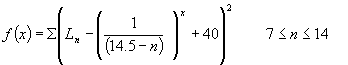|
||||||
| |
||||||
|
The female reproductive cycle is an example
of a real biological oscillating system where the periodic nature of the
system may be described by a set of difference equations. The model is
developed using blood-level of three different ovarian and gonadotropic
hormones, estrogen, progesterone, and luteinizing hormone. We construct
this model using actual sample data describing the levels of these hormones
present in the body throughout the month, then discuss the applications
of such research.
BACKGROUND The female reproductive cycle is an intricate series of chemical secretions and reactions, and delicate changes in physical anatomy, which produce the ultimate potential for fertility and birth. These chemicals, known as hormones, are human proteins with the capacity to affect and regulate human growth and function. The female reproductive cycle is a general term which encompasses the ovarian cycle, the uterine cycle, the hormonal changes that regulate them, and cyclical changes in the breasts and the cervix. The ovarian cycle is the term used to describe the series of events associated with a developing oocyte, or, egg cell, within the ovaries, while the uterine cycle is the preparation of the environment of the uterus for the reception of a fertilized ovum. The regulation of both the ovarian and uterine cycles is the function of several gonadotropic and ovarian hormones. Among the most important of the gonadotropins are luteinizing hormone (LH) and follicle stimulating hormone (FSH), while the most crucial ovarian hormones are the androgens, estrogen and progesterone. The hypothalamus-derived hormone in control of the gonadotropins, and indirectly, all female sexual function, is gonadotropin releasing hormone (GNRH). The release of GNRH in an adolescent female marks the commencement of that female's reproductive capability, and ultimately, the onset of menstruation (Tartora 938-40). Among the most important aspects of the female cycle is the periodic nature in which it occurs. From the time a young woman begins to menstruate, until the time that woman reaches menopause, hormonal pulses and secretions occur extremely regularly, varying little from month to month. This periodicity is the characteristic on which this research capitalizes, since it is the periodicity which we may describe using several mathematical difference equations. These equations are functions of three of the key hormones, luteinizing hormone, estrogen, and progesterone. These hormones were chosen over the rest of the involved hormones for their paramount roles in the regularity of the cycle. It is necessary to understand the biological proceedings which these equations describe. The duration of the female reproductive cycle is typically 24-35 days. This research will assume the average 28-day cycle, in the absence of fertilization and pregnancy, divided into three phases; the menstrual phase, the preovulatory phase and the postovulatory phase. It is the preovulatory phase that is the most variable phase lasting anywhere from 6-13 days and accounts for most of the time difference when cycles are either shorter or longer than the 28-day average, while the postovulatory phase is almost always 14 days in length (Tartora 938). These proceedings begin within the ovaries of a developing female. The ovaries contain oocytes in various stages of development. In the preliminary stages of development an oocyte contains only a single layer of surrounding cells. These cells are referred to as follicle cells. As the egg matures further, and multiple layers of surrounding cells form, the cells are then referred to as granulosa cells. The surrounding cells are responsible for nourishing the developing oocyte, and, as the follicle grows larger, for the production and secretion of estrogen. Estrogen, then, is produced at regular levels throughout childhood. It is during a girl's childhood that GNRH is inhibited from being secreted by the negative feedback effect that estrogen has on the midbrain gland responsible for the output of GNRH, the hypothalamus. As a girl matures toward puberty, the hypothalamus undergoes significant changes in its childhood sensitivity to the ovarian hormone and is stimulated toward the release of GNRH, in a rhythmic, pulse-like manner. As the pulses become stronger and stronger, the release of GNRH into the brain positively effects the anterior pituitary gland to commence the production and secretion of the human gonadotropins, LH and FSH. Continued production of FSH and LH stimulates growth and maturation of the follicles inside the ovaries, and the first preovulatory phase of the female menstrual cycle begins (Marieb 979-81). Since the follicular cells are responsible for the output of estrogen, LH and FSH, in turn, promote estrogen secretion in preparation for ovulation. At this point in the cycle, there are changes that take place within the ovaries. A single primary oocyte is targeted to resume the halted stages of meiosis and develop into a mature follicle, a fluid-filled follicle that will soon rupture and expel a haploid secondary oocyte in the process of ovulation. Maturation of the follicle means increased production of estrogen. This increase in estrogen serum levels exhibits a negative feedback on the anterior pituitary inhibiting the release, but stimulating the production and storage of the gonadotropins. With the cessation of gonadotropin output, estrogen levels continue to rise; and, while the initial small rise in estrogen inhibits gonadotropin release, high estrogen levels have the opposite effect. By the time the ovarian estrogen production reaches its maximum before day 14, the negative feedback effect on the anterior pituitary is reversed to a dramatically positive one and a burst-like surge of stored LH is released (Marieb 982). The LH surge is actually responsible for affecting the final development, and subsequent rupture of the mature follicle. LH also induces the follicle cells to secrete digestive enzymes thereby enabling the secondary oocyte to pass through the ovary wall marking ovulation at day 14 of the cycle (Tartora 940). As the secondary oocyte actually moves through the ovary wall, the outer layer of cells -- the cells responsible for estrogen secretion -- are damaged by the rough contact. Estrogen output is then halted and serum levels drop slowly for a short period immediately following the ovulatory process, and the beginning of the postovulatory phase commences (Marieb 982). The extraordinarily high levels of LH now have another function. Once the secondary oocyte and its remaining surrounding cells are swept into the fallopian tube, luteinizing hormone works to establish the corpus luteum from the remnants of the ruptured mature follicle inside the ovary. The corpus luteum, sometimes referred to as the yellow body, actually stems from a blood clot that develops as the mature follicle ruptures in ovulation. The remaining follicular cells that then enlarge and change character absorb the clot. Under the influence of LH, these enlarged cells become the corpus luteum. The corpus luteum is a gland which maintains the responsibility of both the production and secretion of estrogen and progesterone. As estrogen and progesterone levels again rise, their combined effect on the anterior pituitary's production of the gonadotropins is extremely negative and LH levels decline significantly. Since it is LH that is responsible for the maintenance of the corpus luteum, the marked decline in LH levels also (assuming that the oocyte has not, in the meantime, been fertilized) heralds the degeneration of the corpus luteum into a scar called the corpus albicans. With the corpus luteum no longer producing progesterone and estrogen, the serum levels of the ovarian hormones decline sharply and the menstrual phase may then begin (Tartora 940-1). At this time, the uterine endometrial lining, which had been developing anticipating a pregnancy, is shed in the form of 50-150 ml of blood, mucus, tissue fluid, and epithelial cells. The menstrual flow moves out the uterine cavity to the cervix, then through the vagina to the exterior. The marked decline of the ovarian hormones at the end of the cycle also ends their blockade of the hypothalamic-pituitary axis, thus allowing once again for the release of the gonadotropins (Tartora 939). The cycle then begins anew with the initial climb of LH stimulating the follicles within the ovary to develop toward maturation. After the first menstrual cycle a young woman experiences, the menstrual phase, by convention, marks the beginning of each new cycle. The events of the female reproductive cycle may be easily understood if associated with a timeline. The menstrual cycle lasts roughly five days -- days one through five the average cycle. By day six, the follicle within the ovary is actively secreting estrogen, until estrogen serum levels reach their threshold value at around day 11. Between days 11 and 14 the LH surge is actively promoting oocyte division and follicular rupture so that ovulation may occur at day 14 of the cycle. The corpus luteum is generally established by day 15, and LH levels have already declined by the end of the 16th day. It then takes about 10 days for the corpus luteum to degenerate and estrogen and progesterone to begin their decline at around day 26 of the cycle. In the next two days LH levels are elevated and follicles stimulated to mature so that by the beginning of the new cycle, estrogen levels are once again on the rise (Marieb 982). In summary then, the hormonal cycle may be viewed as the following series of events: 1. Luteinizing hormone rises and stimulates the follicle to produce estrogen 2. As estrogen is produced by the follicle, estrogen levels rise inhibiting the output of LH 3. Estrogen levels continue to rise to the threshold level at which point the negative feedback is reversed and LH surges 4. Ovulation occurs just after the LH surge damaging the estrogen-producing cells which results in a decline in estrogen 5. The LH surge results in the establishment of the corpus luteum -- an estrogen and progesterone secreting gland 6. Estrogen and progesterone serum levels climb steadily, supressing LH output 7. Lack of LH promotes the degeneration of the corpus luteum 8. Cessation of corpus luteum activity means a decline in estrogen and progesterone output 9. The decline in the ovarian hormones ends their negative effect on the secretion of LH 10. LH is secreted and the cycle begins again (Marieb 982).
Knowing these basic trends is paramount
in translating the biological activities associated with the hormonal cycle
into purely mathematical statements a system of difference equations.
After careful analysis of hormone serum levels found in a compilation of
experimental graphs, the decision to use difference equations to describe
the cycle stems from the obvious dependence of each particular daily hormone
serum level on the level of that hormone the previous day. Also of notable
importance is the apparent postovulatory exponential growth of the progesterone
curve, the mid-cycle luteinizing hormone spike, and the seemingly exponential
growth of estrogen in the beginning of the cycle, followed by a second
postovualtory rise concurrent with the rise in progesterone. These general
trends served as a necessary guideline, while care was taken to match equation-generated
values to those determined experimentally. The following set of difference
equations is the product of that quest. The equations, which approximate
the periodicity of the cycle, represent general hormonal trends in terms
of estrogen (E), luteinizing hormone (L), and progesterone (P), where "n"
represents cycle day (Ln = luteinizing hormone level on day
"n").
EQUATIONS AND ANALYSIS (1)
Equation (1) Analysis Realizing that any rise in luteinizing hormone is dependent on whether estrogen is rising or has yet reached its threshold level, there are several terms in equation (1) which accurately account for this relationship. The LH curve demonstrates a minimum of 8 IU/L, also a feature of equation (1). The exponent 1.88 (on (Ln-1 8)1.88 ), describing the rise in luteinizing hormone, was determined by performing on data, non-linear least squares. Values for the luteinizing hormone serum level were taken from a list of experimentally determined serum levels found in an American Fertility Society study on precise measurements of gonadotropins and estrogen at ovulation (Mishell). The non-linear least squares technique involved approximating a minimum value of the following function,
This function was chosen since the steep rise in LH appears to be a function with a horizontal asymptote. Equation (1) has another term relating the level of LH to that of progesterone. This term primarily accounts for the decline in LH as progesterone rises after the ovulation spike, where H(n) (the Heaviside function) is defined as:
The Heaviside function allows for an accurate description of the switch to a positive feedback as estrogen levels climb above their threshold value. In this way, H(n) can be considered to be a functional "on/off switch" for hormones that are similarly affected by the specific level of an accompanying hormone. The next term in equation (1) accounts for the dramatic decrease in LH level immediately following ovulation, where d(n) (the Delta Function) is defined as:
and f+ is defined as:
Estrogen levels rise in the preovualtory
phase according to the first term in equation (2), decline for a short
while, then rise again following the trend in progesterone in the postovulatory
phase. The first term in equation (2) is representative of the approximate
exponential growth displayed by the estrogen serum levels leading up to
ovulation. The decline and subsequent rise is described by the function
g(n), a piecewise function for estrogen serum levels between days 13 and
21. Since hormone serum levels are generally based on biological feedback
mechanisms which are turned on and off as other involved hormones reach
threshold values, a piecewise function of this type was necessary to describe
the trend in estrogen during these days.
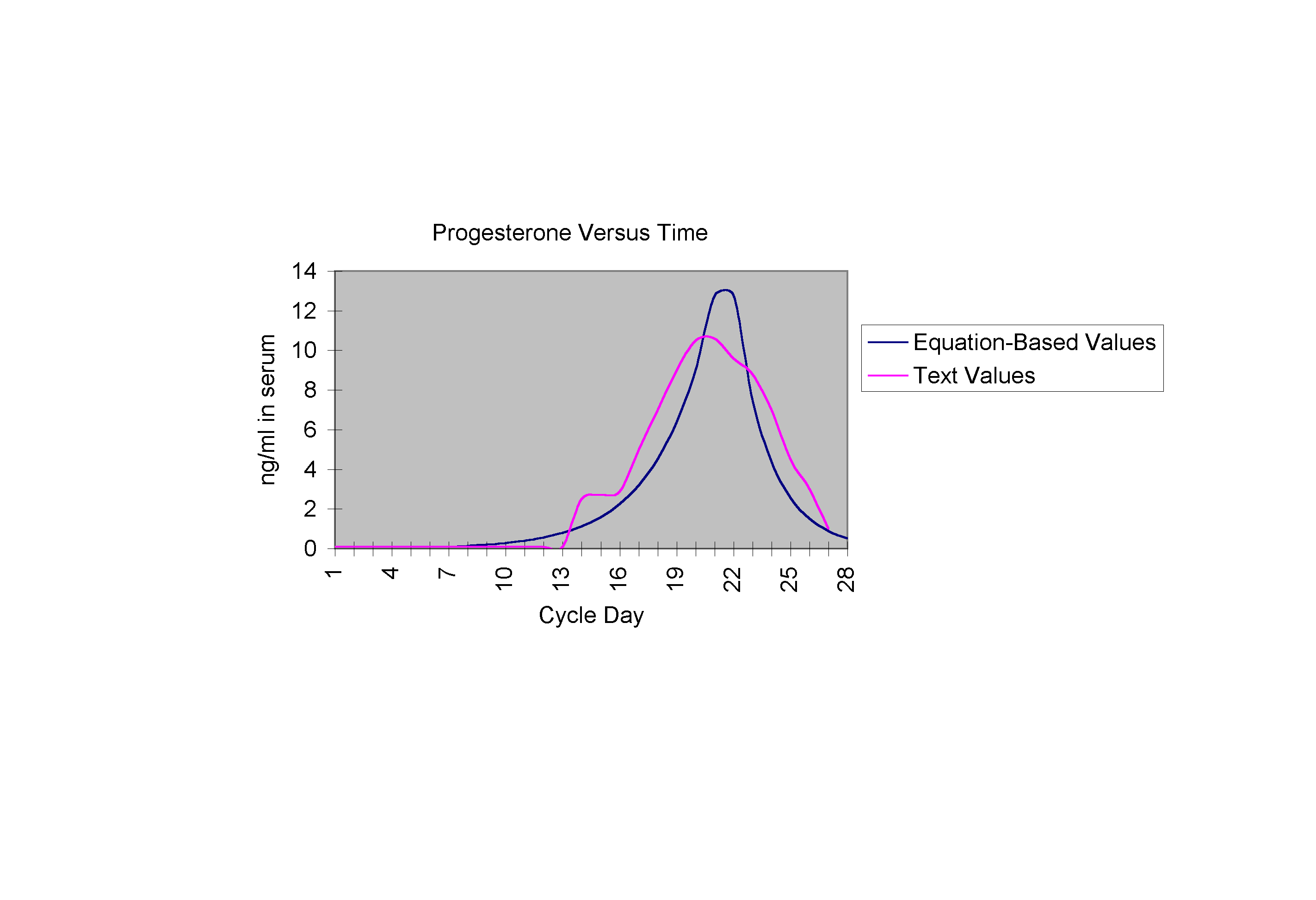
Equation (3) Analysis Equation (3), describing the progesterone level, exhibits a growth that is approximately exponential, then declines with a similar rate just before the end of the cycle. The constant 0.414 reflects this exponential growth, and the function H(n) again maintains the responsibility of switching progesterone growth on and off as necessary.
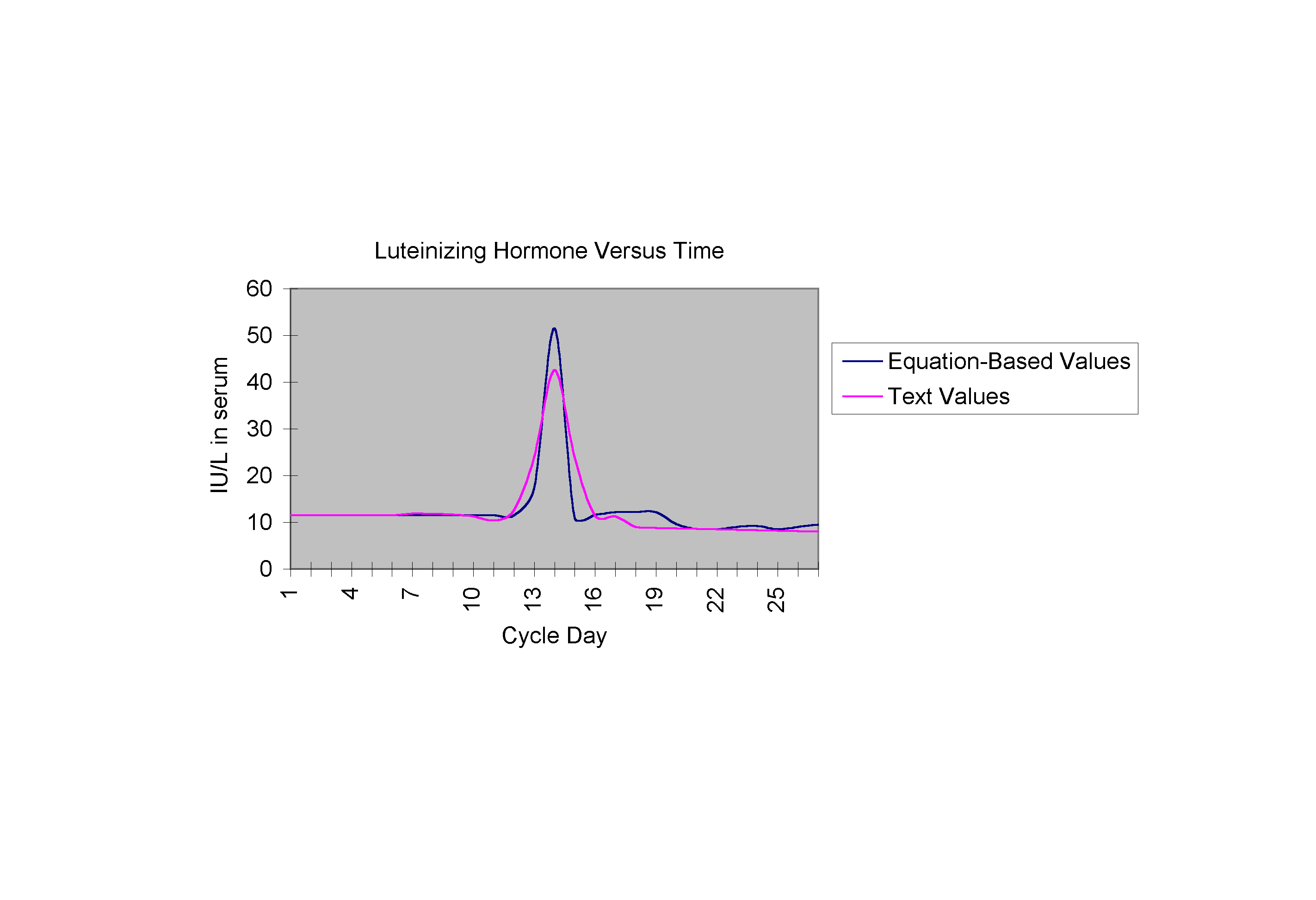
The following graphs are the curves described by equations (1), (2), and (3) respectively, each shown on the same set of axes as the text curve for that hormone (Yen).
CONCLUSION There are obvious strengths in the above equations captured in the trends in the graphs. Graph 1, comparing Ln with actual LH values, demonstrates a mid-cycle spike almost perfectly mirroring the actual curve. Graph 2 demonstrates how closely the growth (both preovulatory and postovulatory) in the estrogen equation mimics that of the actual estrogen values, as does Graph 3 for the progesterone curves. There are also weaknesses in the equations which have to do with the manipulation of terms. Special conditional terms and functions were added to account for several aspects such as the dramatic decrease in LH serum level at day 15, or the postovulatory rise in estrogen which have no direct biological explanations. Further research in this area would, idealistically, be pointed in the direction of reforming these equations so that they specifically reflect the fact that each of the hormone levels are dependent solely upon the level of related hormones and not specific cycle day. These "special terms," thus, would be replaced with terms that make the system not only autonomous, but also non-dimensional. Female reproductive cycle research of this type has many varying practical applications as well. The ability to use precise mathematical statements to predict necessary hormone levels may allow huge advances in the field of endocrinology. Hormone therapy, at present, allows for continued health after the onset of menopause, and also allows for methods in birth control, controlled ovulation, and hormone replacement therapy for hysterectomy patients. Hormone therapy is also now aiding couples who are having difficulty conceiving because of irregular female hormonal cycles, to conceive successfully and give birth to healthy babies. Equations such as the ones presented in this research may allow physicians to better target hormonal inconsistencies, the smallest of which can make ovulation, and thus, conception, impossible. With the ability to make a prediction about the amount of hormone that should be present in an individual at a specific time, treatment plans may be made less general, and more specialized. In the area of cycle regulation, research of this type offers intriguing possibilities.
Best, E.N.: Null space in the Hodgkin-Huxley equations: a critical test. Biophys. J. 27, 87-104 (1979) Filcori M, Butler JP, Crowley WF, Neuroendocrine regulation of the corpus luteum in the human: evidence for pulsatile progesterone secretion. Journal of Clinical Endocrinology and Metabolism. 73:1638 (1984) Hodgkin, A.L., Huxley, A.F.: A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. (London) 117, 500-544 (1952) Hoft JD, Quigley ME, Yen S, Hormonal dynamics at midcycle: a reevaluation. Journal of Clinical Endocrinology and Metabolism. 57: 792 (1983) Marieb, Elaine. Human Anatomy and Physiology. Redwood City: Benjamin/Cummings(1995). Midgly AR Jr. Radioimmunology: A method for human chorionic gonadotropin and human luteinizing hormone. Endocrinology 79:10 (1966) Mishell DR, Davajan V, Lobo RA, Infertility, Contraception, and Reproductive Endocrinology 3rd Ed. London: Blackwell Scientific Publications (1991) Murray, J.D. Mathematical Biology. New York: Springer (1993) Schwan, Herman. Biological Engineering. New York: McGraw-Hill Book Company (1969) Speroff L, Glass RH, Kase NG, Clinical Gynecologic Endocrinology and Infertility 5th ed. Baltimore: Williams and Wilkins (1994) Tartora, Gerard and Sandra Grabowski. Principles of Anatomy and Physiology. New York: HarperCollins(1996). Vermesh M, Kletzky OA, Longitudinal evaluation of the luteal phase and its transition into the follicular phase. Journal of Clinical Endocrinology and Metabolism. 65: 653 (1987) Winfree, A.T.: Resetting Biological Clocks. Physics Today 28, 34-39 (1975) Yen S, Jaffe R, Reproductiove Endocrinology 3rd Ed., Physiology, Pathophysiology, and Clinical Management. Philadelphia: Harcourt, Brace, Jovanovich Inc. (1991)
|
|||||||