|
||||||
| |
||||||
|
Name: Elizabeth Betty Gilpin Present Year in Education: Senior E-mail Address:Staupin@ix.netcom.com Candidate for degree? Yes
Have you identified a research mentor for
a senior thesis (if applicable)? Yes
PROJECT SUMMARY The purpose of this study is to investigate the effects of nutrient availability on epibiont (attached plants and animals) abundance and diversity in seagrass communities within the estuaries of southwest Florida. Research will be conducted in the Northern Charlotte Harbor estuary system. It is expected that epibiont abundance will be directly correlated with nutrient availability, and plant and animal epibiont ratios will be related to water quality. Two environmental locations will be examined: dark-colored, inner estuary waters near Punta Gorda, Florida; and lighter more saline waters near Boca Grande, Florida. These locations have very different water quality characteristics. The Punta Gorda location is highly influenced by freshwater inflow; it receives more detrital matter than the Boca Grande site, and the salinity range is significantly lower. These locations have been chosen because of the differences in their water quality characteristics and their location within the estuarine system. Two turtle grass (Thalassia testudinum) beds will be examined at each location. One bed at each setting will be located adjacent to urban storm water runoff. The other site will be located in a relatively un-impacted area and serve as a control. Seagrass shoots will be collected at each of the four T. testudinum beds, epibionts will be scraped, and epibiont abundance will be determined per shoot. Seagrass nutrient availability will be determined through seagrass tissue analysis. Plant and animal epibiont ratios will be determined through epiphyte chlorophyll concentrations. Each bed will also be analyzed for water column nutrient concentrations. Results from this project will help to
determine if epibiont abundance can provide an early indication of increased
nutrient enrichment, changes in nutrient availability, and ecosystem decline.
Cover Sheet.............................................................................i
PROJECT DESCRIPTION Statement of the Problem Seagrass meadows are the aquatic equivalent of terrestrial tropical rain forests. Seagrasses are extremely productive ecosystems, providing food, shelter, and nursery facilities for many marine organisms. Unfortunately, however, seagrasses worldwide are declining at alarming rates. In Florida, Tampa Bay has lost approximately 80% of its seagrass cover from historic levels (Zieman and Zieman 1989); Sarasota Bay has lost about 30% of its seagrasses since the 1940s (Mangrove Systems, Inc. 1988); and Charlotte Harbor, the area of focus in this study, has lost approximately 29% of its seagrasses from historical levels (Duke and Kruczynski 1992). Loss of seagrasses around the world has
most often been linked to anthropogenic (human induced) nutrient enrichment
of nearshore waters (Larkum 1976; Kemp et al. 1983; Cambridge and McComb
1984; Orth and Moore 1984; Silberstein et al. 1986; Neverauskas 1987; Valiela
et al. 1990; Reyes and Merino 1991; Tomasko and Lapointe 1991; Lapointe
et al. 1994). One of the mechanisms towards this loss is overgrowth of
seagrasses by epibionts (figure 1) in areas of enhanced nutrient availability,
which in turn reduces seagrass photosynthesis and growth rate (Phillips
et al. 1978; Kemp et al. 1983; Cambridge et al.1986; Frankovich and Fourqurean
1997).
Traditionally, increased water column nutrients, enhanced epibiont levels, and the decline in seagrass health were seen as evidence of anthropogenic nutrient enrichment (Cambrige and McComb 1984; Siberstein et al. 1986; Tomasko and Lapointe 1991). More recent studies have demonstrated that these traditional indicators do not fully describe nutrient availability within an estuary (Neckles et al. 1993; Tomasko et al. 1996). Seagrasses thrive in estuarine areas of good water clarity with low nutrient enrichment and low nutrient availability. When nutrients are introduced into the system from increased pollutant loads and surface water runoff, nutrient availability is enhanced, creating conditions favorable for the establishment of seagrass epibionts and water column algae. The active uptake of nutrient by epibionts and algae, and the top down grazing of higher level organisms, leads to an eventual net loss in nutrient availability for seagrasses, resulting in stress and decline in seagrass communities (Frankovich and Fourqurean 1997). This active uptake of increased nutrients keeps water column nutrient concentrations low, and the grazing control of epibionts may actually hide the effects of nutrient enrichment (Tomasko et al. 1996). Much of the research within the last ten years has associated anthropogenic nutrient enrichment of coastal marine waters with the development of plant epibionts or epiphytes on the leaves of various species of seagrasses (e.g. Kemp et al. 1983; Borum 1985; Twilley et al.1985; Williams and Ruckelshaus, 1993; Neckles et al. 1993; Coleman and Burkholder 1994). Some authors have suggested that epiphyte biomass may be a useful indicator of eutrophication (Borum 1985; Tomasko and Lapointe 1991; Tomasko et al., 1996), while others have emphasized the positive role of epiphytes on aquatic systems in terms of primary production, nutrient exchange, and food production (Harlin 1973; McRoy and Goering 1974; Penhale 1977; Penhale and Thayer 1980). Despite the recent interest in epiphytes, there have been few quantitative studies linking epizoans to increased epiphytes, nutrient enrichment or nutrient availability. Epizoan carbonate, however, can account for as much as 70 to 80% of the total epibiont standing stock (Heijs 1984; Frankovich and Zieman 1994). The general pattern of distribution of plant and animal epibionts is from a dominance of epizoans in areas of low salinity and dark waters, to a gradual shift in dominance of epiphytes in lighter marine waters (Tomasko 1998). This pattern of distribution is extremely important when considering Charlotte Harbor, Florida. Northeastern Charlotte Harbor receives fresh water from three major tributaries: the Myakka River and Peace River to the North, and the Caloosahatchee River to the East. However, the western portion of Charlotte Harbor, along the Gulf of Mexico, becomes almost entirely marine towards the gulf passes. One would therefore expect to find a shift in plant/animal communities along an east to west gradient within the harbor. The purpose of this study is to further
our understanding concerning the effects of nutrient availability on epibiont
biomass and the patterns of plant and animal epibiont distribution.
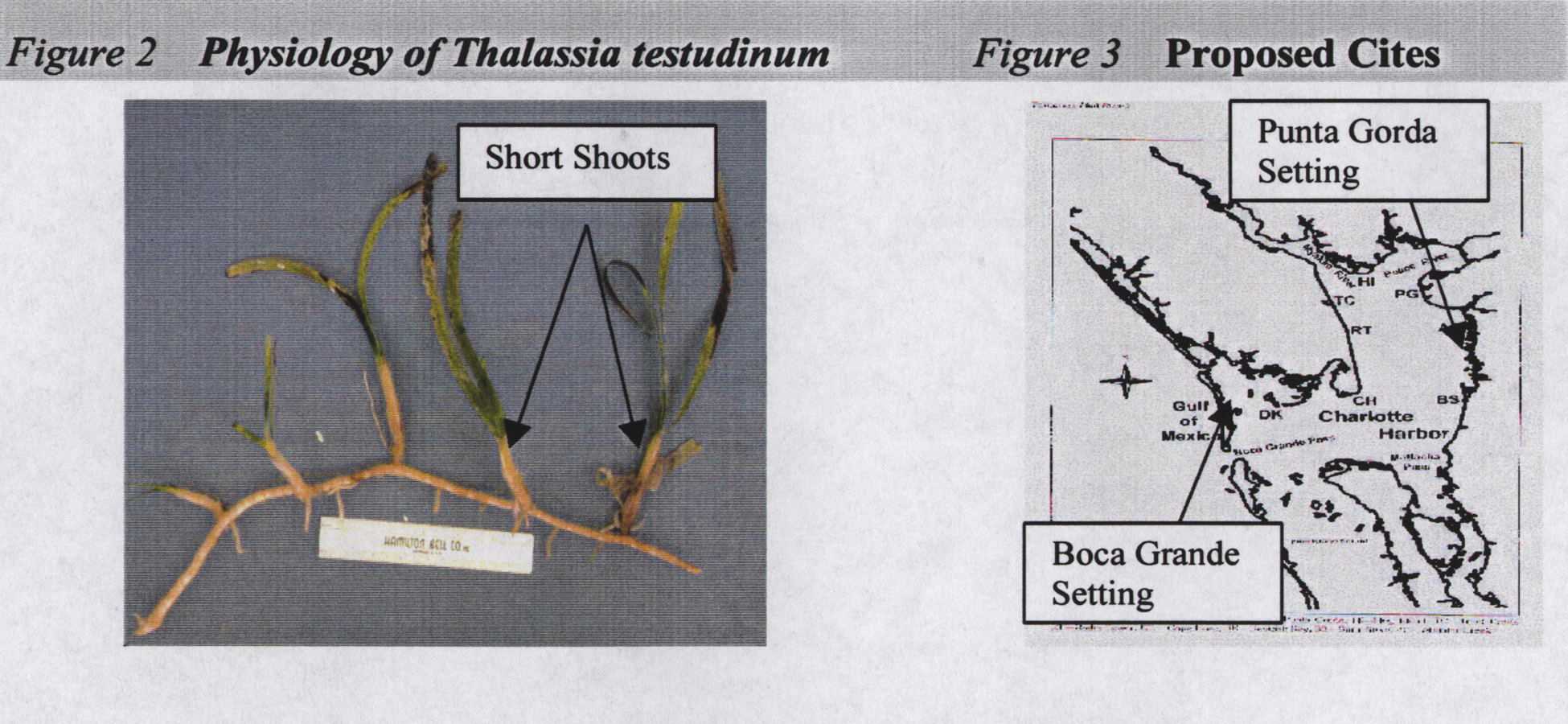
Research will include comparing epibiont loads, seagrass short shoot size
(figure 2), nutrient availability to seagrasses, specifically Thalassia
testudinum, and water column nutrient concentrations in two Northern
Charlotte Harbor estuarine settings. This study will test two hypotheses:
Research Objectives Two estuarine settings will be investigated: 1.) the fresh, dark, tannin waters off Punta Gorda in northeastern Charlotte Harbor; and 2.) the lighter, marine waters off Boca Grande in western Charlotte Harbor (figure 3). Settings have been chosen due to the high degree in variance with respect to salinity and water quality. These differences should provide a clear relationship between water quality including salinity and plant/animal ratios. Two Thalassia testudinum beds will be identified at each of the settings. Each setting will contain one bed adjacent to urban storm water runoff, and one bed in a primarily undisturbed area. In order to eliminate the possibility of bias, careful consideration will be taken to ensure all confounding variables are accounted for. The Punta Gorda and Boca Grande settings will have similar water depth and seagrass populations, and within each setting, test and control sites will have similar salinities and water color. Once seagrass beds have been identified, they will be monitored bi-weekly for a period of two months. Patterns of benthic nutrient availability, indicated by seagrass tissue nutrient concentrations of nitrogen and phosphorus, will be compared to water column nutrient concentrations. In addition, total epibiont load, epiphyte chlorophyll load, and seagrass short shoot size will be examined. Methods Study Site Four Thalassia testudinum beds will be identified in northern Charlotte Harbor. Each bed will be marked with a buoy, coordinates identified using a GPS, and physical landmarks noted. Physical parameters (salinity, pH, dissolved oxygen) of each bed will be characterized using a YSI or Hydrolab data sonde. Light availability will be measured using a secchi disk, and water color and turbidity will be determined by using the Platinum Colbalt method and nephthelometry respectively (Greenberg et al. 1992). Seagrass & Epibiont Sampling Three T. testudinum short shoot samples (figure 2) will be collected at each of the four beds by randomly enumerating 10 short shoots in replicate 25cm2 quadrates. Epibionts will be removed from the collected seagrass leaves by gently scraping with a double-edged razor (Frankovich & Zieman 1995). Both seagrass short shoots and epibionts will be dried, then total epibiont load (mg dry wt of epibionts / g dry wt of scraped short shoot) will be determined from the epibiont dry weight and the dry weight of the scraped short shoots. In order to determine the ratio of floral epibionts to faunal epibionts, epiphytes will be quantified by extracting chlorophyll a from the separated epibiont material scraped from each short shoot. Chlorophyll a concentrations will be determined using spectrophotometry (Greenberg et al. 1992). Epiphyte chlorophyll loads will be determined based on the chlorophyll a concentrations and the dry weight of the scraped short shoots (ug chl a / mg short shoot). In order to determine differences in plant versus animal epibionts, the autotrophic index AI=ug chl a mg-1 epibiont, will be used (Greenberg et al. 1992). The availability of nutrients to the seagrasses will be determined by analyzing the carbon, nitrogen, and phosphorus content of the seagrass leaves (Fourqurean et al. 1992a, b). Carbon (C) and nitrogen (N) contents will be determined by Nitrogen/Carbon Analyzer (Carlo Erba, NA 1500). Phosphorus (P) concentrations will be examined after dry oxidation and acid hydrolysis using methods as described by Solorzano and Sharp (1980). Water Quality During each seagrass sampling event, water quality parameters will be collected for comparison with epibiont loads and nutrient content of seagrasses. Water quality parameters to be analyzed include: total organic carbon (TOC), total organic nitrogen (TON), ammonium (NH4+), nitrate (NO3-), nitrite (NO2-), and total phosphorus (TP), as well as water column chlorophyll a, salinity, color, and turbidity. Nutrient samples will be collected in 500 ml I-Chem bottles, preserved with H2SO4, downstream along a transect adjacent to the point source and an equal distance from each bed. Samples will be analyzed using a Technicon Autoanalyzer II according to Technicon (1973). Chlorophyll a, water color, and turbidity samples will be collected in a 1L Nalgene light resistant bottle, immediately placed in an ice slurry. Chlorophyll a concentrations will be determined using spectrophotometry (Greenberg et al. 1992). Turbidity will be measured using nephthelometry (Greenberg et al. 1992), and water color will be visually determined using the Platinum Colbalt method (Greenberg et al. 1992). Salinity will be measured using a YSI or Hydrolab. Statistical Analysis Before statistical analysis is conducted, all data will be examined for normality and where appropriate data will be ln-transformed. Since the ecological balance of the Charlotte Harbor estuarine system is nitrogen limited, and nutrient inputs are inversely correlated to the salinity of inflow waters, total nitrogen (TN) vs. salinity and total inorganic nitrogen (TIN) vs. salinity will be plotted to determine the rates of nitrogen uptake by seagrasses from the water column. Comparisons of the responses of epibionts to nutrient availability will be assessed using one and two way analysis of variance (ANOVA) to detect differences in mean epibiont abundance between sites and dates. Comparisons between seagrass biomass and nitrogen availability will be made using correlation analysis. Expected Results and Significance Experimental results are expected to indicate that water column nutrients and benthic nutrient availability are correlated to the distribution of epibiont communities on T. testudinum in Charlotte Harbor, Florida. It is also predicted that the ratio of faunal epibionts to floral epibionts will be higher in the eastern Charlotte Harbor settings where the waters are more tidally-restricted and more heavily influenced by fresh water inflows. This will be determined by the epiphyte chlorophyll loads. Time Table and Project Management This project will begin July 2000 and last two months. Parameters will be monitored and analyzed bi-weekly. Primary research will be conducted by Betty Gilpin with the help of at least one volunteer. Laboratory analysis will be carried out at a Department of Health certified facility. Ethical Considerations Seagrasses in Charlotte Harbor are the bases for the biological diversity and balance of the estuarine ecosystem. Seagrasses should be protected from future degradation or loss in order to maintain the long term estuary health. As Floridas coastal communities become more heavily populated, stormwater runoff increases, increasing the likelihood of elevated nutrient input. Also, many existing coastal residences are older and have inadequate septic systems. These facts will have to be weighed carefully when making future land use management decisions. Protecting the seagrasses by implementing stricter runoff regulations should be in balance with the rights of individuals who live adjacent to the harbor. REFERENCES CITED Borum J 1985. Development of epiphytic communities on eelgrass (Zotera marina) along a nutrient gradient in a Danish estuary. Mar Biol. 87:211-218. Cambridge ML, Chiffings AW, Brittan C, Moore L, McComb AJ. 1986. The loss of seagrass in Cockburn Sound, Western Australia. II. Possible causes of seagrass decline. Aquatic Botany. 24:269-285. Cambridge ML, McComb AJ. 1984. The loss of seagrass in Cockburn Sound, Western Australia. I. The time course and magnitude of seagrass decline in relation to industrial developments. Aquatic Botany. 20:229-243. Coleman VL, Burkholder JM. 1994. Community structure and productivity of epiphytic microalgae on eelgrass (Zostera Marina L.) under water-column nitrate enrichment. J. Exp Mar Biol Ecol. 179:29-48. Duke TD, Krucynski WL. 1992. A summary of the report: Status and trends of emergent and submerged vegetated haabitats of the Gulf of Mexico, USA, p. 11-28. In The Environmental and Economic Status of the Gulf of Mexico. Gulf of Mexico Program, Stennis, Mississippi. Frankovich TA, Fourqurean JW. 1997. Seagrass epiphyte loads along a nutrient availability gradient, Florida Bay, USA. Mar Ecol Prog Ser. 159:37-50. Frankovich TA, Zieman JC. 1994. Total epiphyte and epiphytic carbonate production on Thalassia testudinum across Florida Bay. Bull Mar Sci. 54:679-695. Greenberg AE, Clesceri LS, Eaton AD. eds. 1992. Standard Methods for Examination of Water and Wastewater. 18th ed. American Public Health Association. 2(1-10), 10(17-19). Harlin MM. 1980. Seagrass epiphytes. In: Phillips RC, McRoy CP. eds. Handbook of seagrass biology: an ecosystem perspective. Garland STPM Press, New York. p 117-131. Heijs FML. 1984. Annual biomass and production of epiphytes in three monospecific seagrass communities of Thalassia hemprichii (Ehrenb.) Aschers Aquat Bot. 20:195-218. Kemp WM, Boynton WR, Twilley RR, Stevenson JC, Means JC. 1983. The decline of submerged vascular plants in upper Chesapeake Bay: Summary of results concerning possible causes. Mar Techn Soc Jour. 17:78-79. Lapointe BE, Tomasko DA, Matzie W. 1994. Trophic structuring of marine plant communities in the Florida Keys. Bull Mar Sci. 54:696-717. Larkum AWD. 1976. Ecology of Botany Bay. I. Growth of Posidonia australis (Brown) Hook. f. in Botany Bay and other bays of the Sydnew Basin. Australian J of Mar and Freshwater Res. 27:117-127. McRoy CP, Goering JJ. 1974. Nutrient transfer between the seagrass Zostera Marina and its epiphytes. Nature. 248:173-174. Neckles, Wetzel RL, Orth RJ. 1993. Relative effects of nutrient enrichment and grazing on qpiphyte-macrophyte (Zostera marina L.) dynamics. Oecologia. 93:285-295. Penhale PA. 1977. Macrophyte-epiphyte biomass and productivity in an eelgrass (Zostera marina L.) community. J Exp Mar Biol Ecol. 26:211-224. Penhale PA, Thayer GW. 1980. Uptake and transfer of carbon and phosphorus by eelgrass (Zostera marina) and its epiphytes. J Exp Mar Biol Ecol. 42:113-123. Phillips GL, Eminson D, Moss B. 1978. A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters. Aquat Bot. 4:103-126. Savarese M. 1998. pers. comm. Florida Gulf Coast University, Ft. Myers, Florida. Silberstein K, Chiffings AW, McComb AJ. 1986. The loss of seagrass in Cockburn Sound, Western Australia. III. The effect of epiphytes on productivity of Posidonia australis Hook. F. aquatic Botany. 24:355-371. Tomasko DA. 1998. Pers Comm. Southwest Florida Water Management District, Tampa, Florida. Tomasko DA, Dawes CJ, Hall MO. 1996. the effects of anthropogenic nutrient enrichment on turtle grass (Thalassia testudinum) in Sarasota Bay, Florida. Estuaries. 19:448-456. Tomasko DA, Lapointe BE. 1991. Productivity and biomass of Thalassia testudinum as related to water column nutrient availability and epiphyte levels: field and Environmental Studies. Mar Ecol Prog Ser. 75:9-16. Twilley RR, Kemp WM, Staver KW, Stevenson JC, Boynton WR. 1985. Nutrient enrichment of estuarine submersed vascular plant communities. 1. Algal growth and effects on production of plants and associated communities. Mar Ecol Prog Ser. 23:179-191. Valiela I, Costa J, Foreman K, Teal JM,
Howes B, Aubrey D. 1990. Transport of groundwater-bourn
nutrients from watersheds and their effects on coastal waters.
Biogeochemistry. 10:177-197.
Zieman JC, Aieman RT. 1989.
The ecology of the seagrass meadow of the west coast of Florida:
a community profile. U.S. Fish and Wildlife Service, Biological Report
85 (7.25).
Contact Information
Education 1996 - A.A., Biological Sciences, Edison Community College, Ft. Myers, Florida 1998 - B.A., Environmental Sciences (in progress), Florida Gulf Coast University, Ft. Myers, Florida
1997 - Present Water Quality Monitoring Assistant, Florida Department of Environmental Protection, Punta Gorda, Florida 1998 - Present Project Coordinator, Charlotte Harbor Estuaries Volunteer Water Quality Monitoring Network 1998 - Present Volunteer, Southwest
Florida Water Management District, Ambient Seagrass
Study
EXPECTED EQUIPMENT AND SPECIAL RESOURCE
NEEDS
|
|||||||